| Cat. # | Size | Qty. | Price |
|---|---|---|---|
| 85493T | 1 Kit (9 x 20 microliters) |
|
| Product Includes | Quantity | Applications | Reactivity | MW(kDa) | Isotype |
|---|---|---|---|---|---|
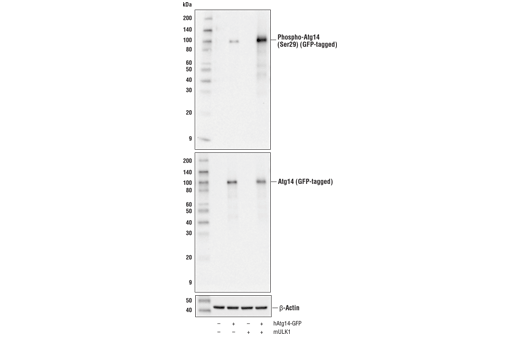
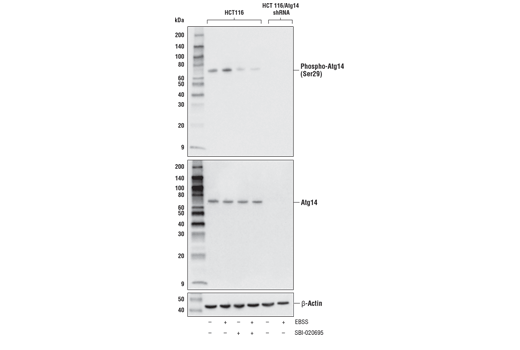
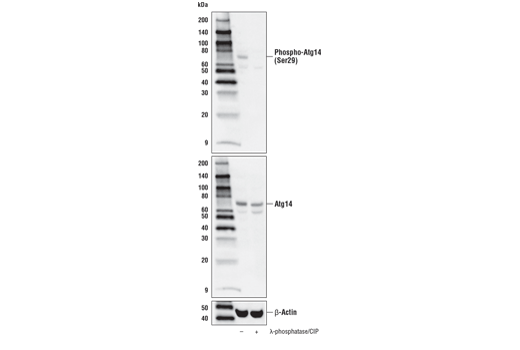
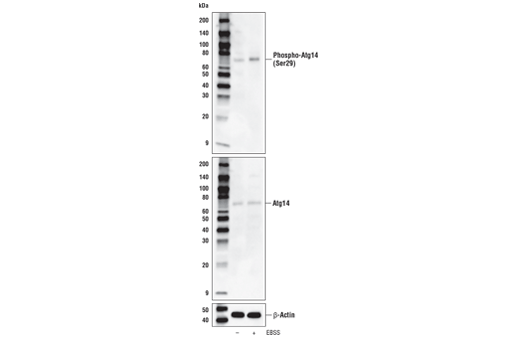
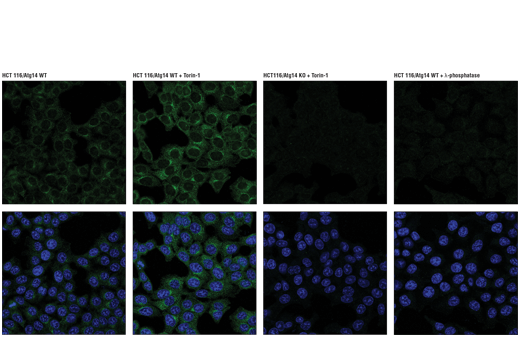
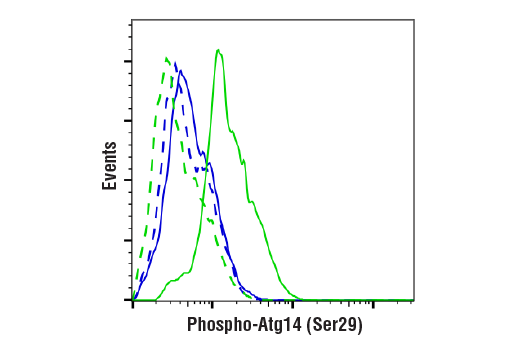
| Phospho-Atg14 (Ser29) (D4B8M) Rabbit mAb 92340 | 20 µl |
|
H M R | 65 | Rabbit IgG |
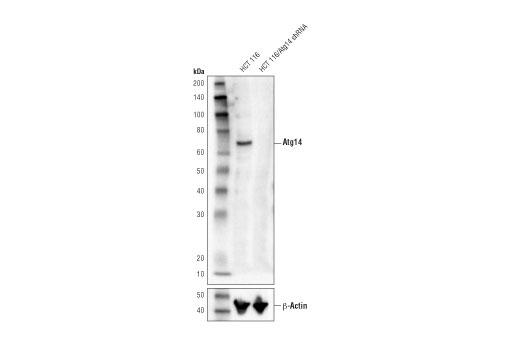
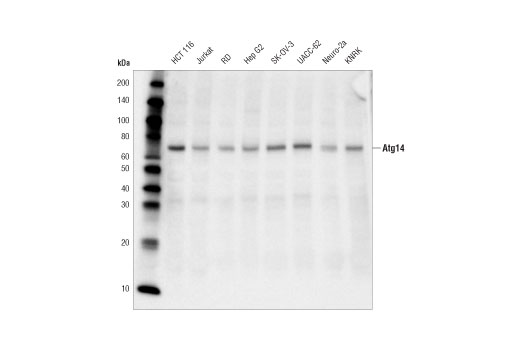
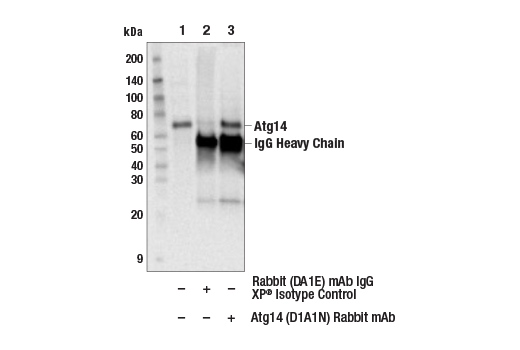
| Atg14 (D1A1N) Rabbit mAb 96752 | 20 µl |
|
H M R | 65 | Rabbit IgG |
| Phospho-Beclin-1 (Ser15) (D4B7R) Rabbit mAb 84966 | 20 µl |
|
H M | 60 | Rabbit IgG |
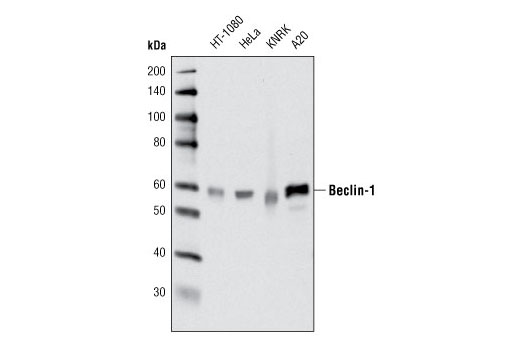
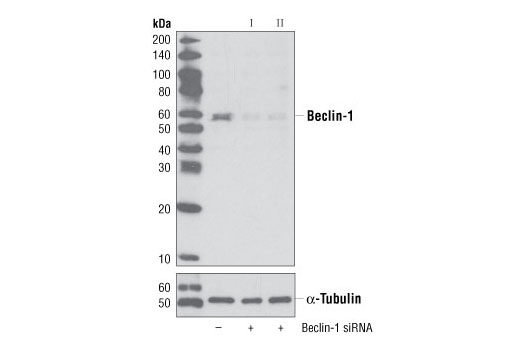
| Beclin-1 (D40C5) Rabbit mAb 3495 | 20 µl |
|
H M R Mk | 60 | Rabbit IgG |
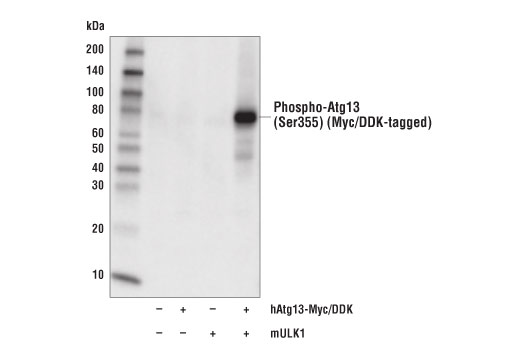
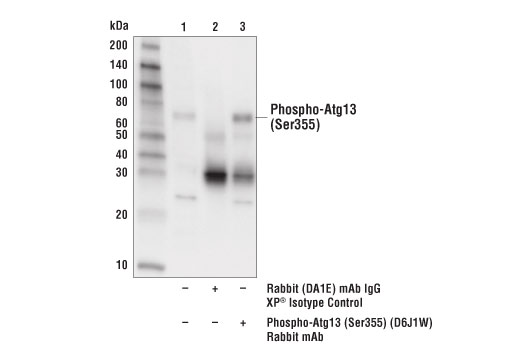
| Phospho-Atg13 (Ser355) (D6J1W) Rabbit mAb 26839 | 20 µl |
|
H | 72 | Rabbit IgG |
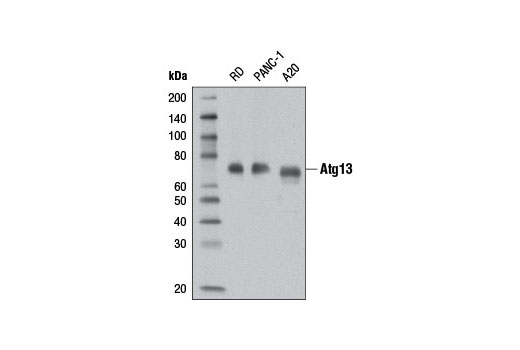
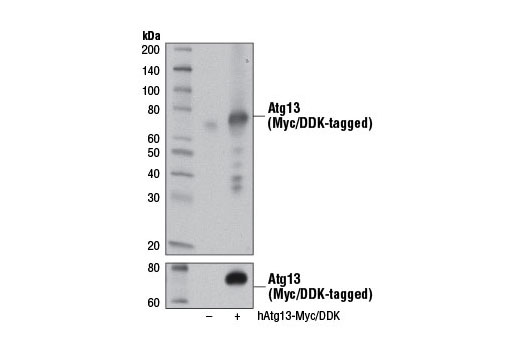
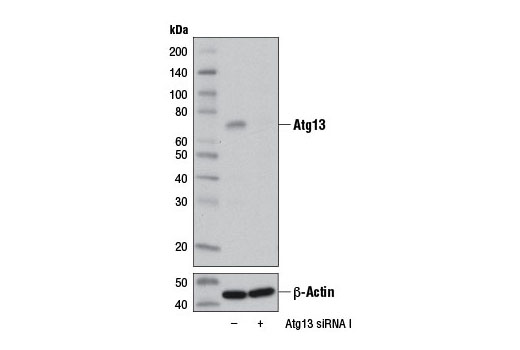
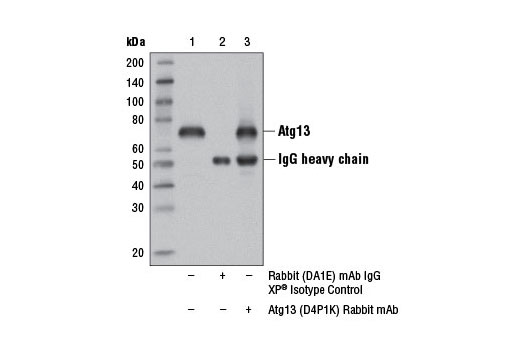
| Atg13 (D4P1K) Rabbit mAb 13273 | 20 µl |
|
H M R | 72 | Rabbit IgG |
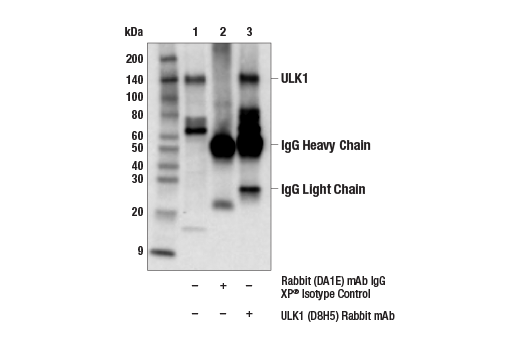
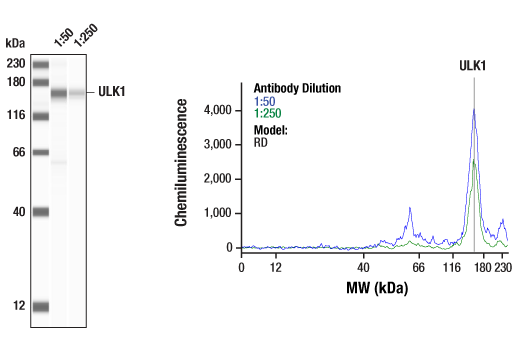
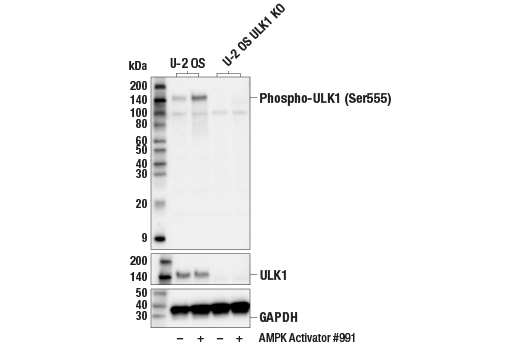
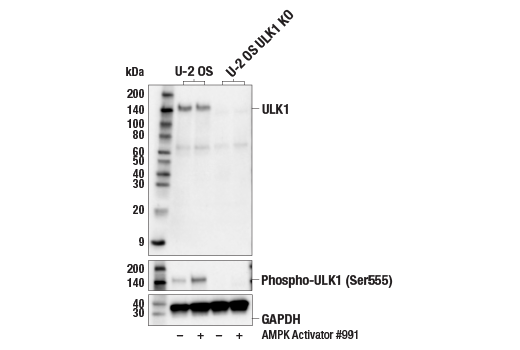
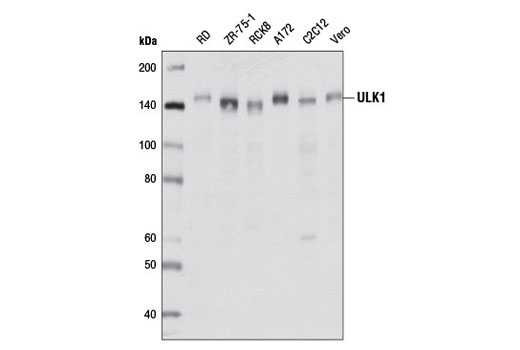
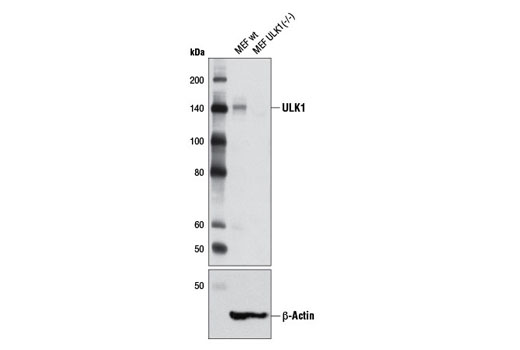
| ULK1 (D8H5) Rabbit mAb 8054 | 20 µl |
|
H M R Mk | 150 | Rabbit IgG |
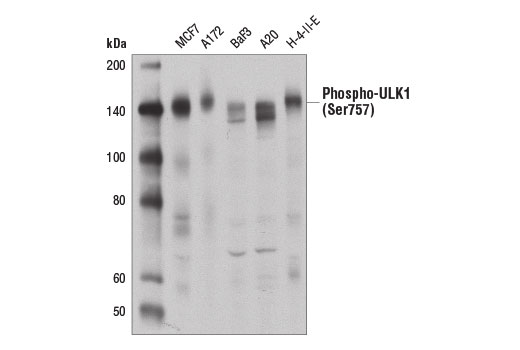
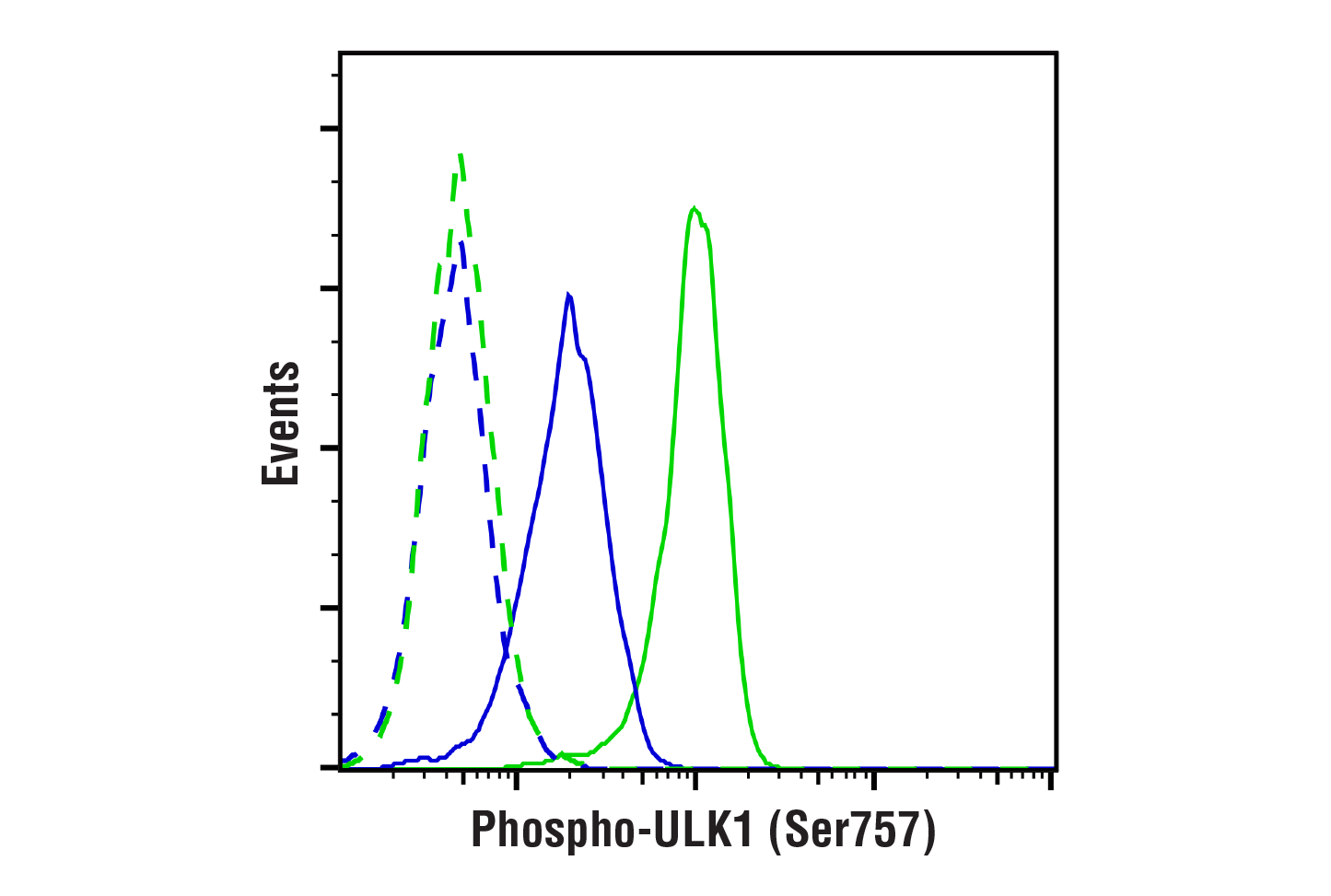
| Phospho-ULK1 (Ser757) (D7O6U) Rabbit mAb 14202 | 20 µl |
|
H M R Mk | 140-150 | Rabbit IgG |
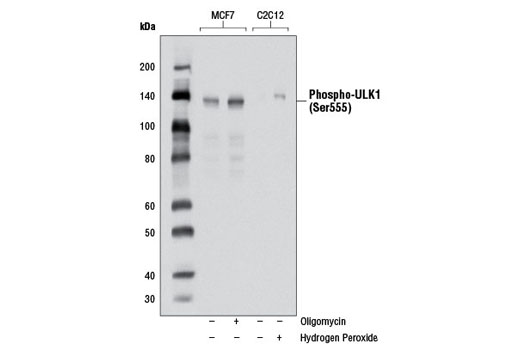
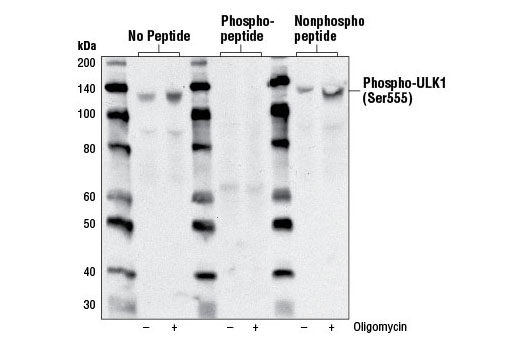
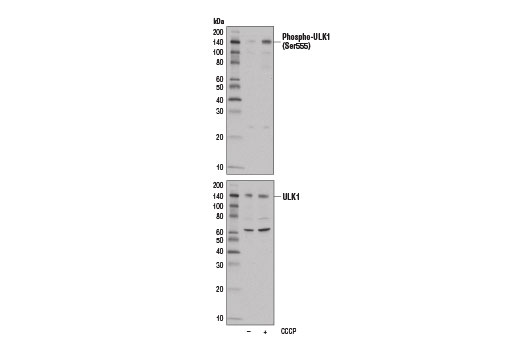
| Phospho-ULK1 (Ser555) (D1H4) Rabbit mAb 5869 | 20 µl |
|
H M | 140-150 | Rabbit IgG |
| Anti-rabbit IgG, HRP-linked Antibody 7074 | 100 µl |
|
Rab | Goat |
Product Information
Monoclonal antibodies are produced by immunizing rabbits with synthetic peptides corresponding to residues surrounding Arg70 of human Atg14, Thr72 of human Beclin-1, Asp462 of human Atg13, and Arg600 of human ULK1. Phosphorylation-specific monoclonal antibodies are produced by immunizing rabbits with synthetic phospho-peptides corresponding to Ser29 of Atg14, Ser15 of human Beclin-1, Ser355 of human Atg13 (Ser318 of isoform 2 of Atg13), Ser555 of mouse ULK1 (equivalent to Ser556 of human ULK1), and Ser757 of mouse ULK1 (equivalent to Ser758 of human ULK1).
Two related serine/threonine kinases, UNC-51-like kinase 1 and 2 (ULK1, ULK2), were discovered as mammalian homologs of the C. elegans gene unc-51 in which mutants exhibited abnormal axonal extension and growth (1-4). Both proteins are widely expressed and contain an amino-terminal kinase domain followed by a central proline/serine rich domain and a highly conserved carboxy-terminal domain. The roles of ULK1 and ULK2 in axon growth have been linked to studies showing that the kinases are localized to neuronal growth cones and are involved in endocytosis of critical growth factors, such as NGF (5). Yeast two-hybrid studies found ULK1/2 associated with modulators of the endocytic pathway, SynGAP, and syntenin (6). Structural similarity of ULK1/2 has also been recognized with the yeast autophagy protein Atg1/Apg1 (7). Knockdown experiments using siRNA demonstrated that ULK1 is essential for autophagy (8), a catabolic process for the degradation of bulk cytoplasmic contents (9,10). It appears that Atg1/ULK1 can act as a convergence point for multiple signals that control autophagy (11), and can bind to several autophagy-related (Atg) proteins, regulating phosphorylation states and protein trafficking (12-16).~AMPK, activated during low nutrient conditions, directly phosphorylates ULK1 at multiple sites, including Ser317, Ser555, and Ser777 (17,18). Conversely, mTOR, which is a regulator of cell growth and is an inhibitor of autophagy, phosphorylates ULK1 at Ser757 and disrupts the interaction between ULK1 and AMPK (17). ULK1 has been shown to phoshorylate several targets in the autophagy pathway, including Ser29 of Atg14, Ser15 of Beclin-1, and Ser355 of Atg13 (19-22).
Explore pathways related to this product.
STRING - Known and Predicted Protein-Protein Interactions.
Except as otherwise expressly agreed in a writing signed by a legally authorized representative of CST, the following terms apply to Products provided by CST, its affiliates or its distributors. Any Customer's terms and conditions that are in addition to, or different from, those contained herein, unless separately accepted in writing by a legally authorized representative of CST, are rejected and are of no force or effect.
Products are labeled with For Research Use Only or a similar labeling statement and have not been approved, cleared, or licensed by the FDA or other regulatory foreign or domestic entity, for any purpose. Customer shall not use any Product for any diagnostic or therapeutic purpose, or otherwise in any manner that conflicts with its labeling statement. Products sold or licensed by CST are provided for Customer as the end-user and solely for research and development uses. Any use of Product for diagnostic, prophylactic or therapeutic purposes, or any purchase of Product for resale (alone or as a component) or other commercial purpose, requires a separate license from CST. Customer shall (a) not sell, license, loan, donate or otherwise transfer or make available any Product to any third party, whether alone or in combination with other materials, or use the Products to manufacture any commercial products, (b) not copy, modify, reverse engineer, decompile, disassemble or otherwise attempt to discover the underlying structure or technology of the Products, or use the Products for the purpose of developing any products or services that would compete with CST products or services, (c) not alter or remove from the Products any trademarks, trade names, logos, patent or copyright notices or markings, (d) use the Products solely in accordance with CST Product Terms of Sale and any applicable documentation, and (e) comply with any license, terms of service or similar agreement with respect to any third party products or services used by Customer in connection with the Products.