Recombinant antibodies offer several key advantages compared to traditional antibodies. These include superior lot-to-lot consistency, continuous supply, and animal-free manufacturing. As such, recombinant antibodies are seeing increased use for scientific research, especially as a means of addressing the ongoing reproducibility crisis.
Traditional polyclonal and monoclonal antibodies are the product of normal B cell development and genetic recombination. They are generated by immunizing an animal with an antigen to elicit an immune response. While polyclonal antibodies are secreted by many different B cell clones and recognize multiple antigenic epitopes, monoclonals originate from a single B cell clone and are specific for just one epitope.
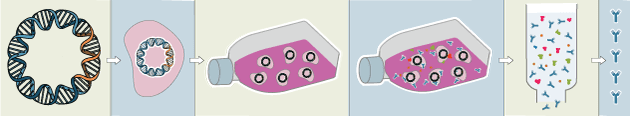
Recombinant antibodies are monoclonal, but their production involves in vitro genetic manipulation. After cloning the antibody genes into an expression vector, this is then transfected into an appropriate host cell line for antibody expression. Mammalian cell lines are most commonly used for recombinant antibody production, although cell lines of bacterial, yeast, or insect origin are also suitable.
Because recombinant antibody production involves sequencing the antibody light and heavy chains, it is a highly controlled and reliable process. In contrast, hybridoma-based systems for producing monoclonal antibodies are subject to genetic drift and instability, increasing the potential for lot-to-lot variability or loss of antibody expression. Recombinant antibodies are highly consistent from lot to lot, thereby ensuring reproducible experimental results.
In vitro methods for producing antibodies are amenable to large-scale production, meaning antibody availability is unlikely to become a limiting factor. Moreover, since the recombinant antibody sequence is known, continuity of supply is assured; in situations where an antibody will be used to support large, long-term studies, this can be an especially critical factor.
Unlike traditional methods for antibody production, recombinant approaches avoid the need to use animals. Where polyclonal antibodies are purified directly from the serum of the immunized host, and monoclonals are purified from either hybridoma-derived tissue culture supernatant or ascites, recombinant antibodies are instead purified from the tissue culture supernatants of transfected host cell lines. Regardless of whether an antibody is polyclonal, monoclonal or recombinant, it must always be properly validated in the intended application prior to experimental use. At CST, we adhere to the Hallmarks of Antibody Validation™, six complementary strategies for determining the specificity, sensitivity, and functionality of an antibody in any given assay. By carefully tailoring these strategies to each antibody product, we guarantee that CST antibodies will work as expected, to help you achieve results you can trust.

| Cat. # | Size | Qty. | Price |
|---|---|---|---|
| 74281SF | 100 µg |
|
| REACTIVITY | H M R Hm Mk Dm Sc |
| SENSITIVITY | Endogenous |
| MW (kDa) | 62 |
| Source/Isotype | Rabbit IgG |
Product Information
Human, Mouse, Rat, Hamster, Monkey, D. melanogaster, S. cerevisiae
Chicken, Zebrafish, Bovine, Pig
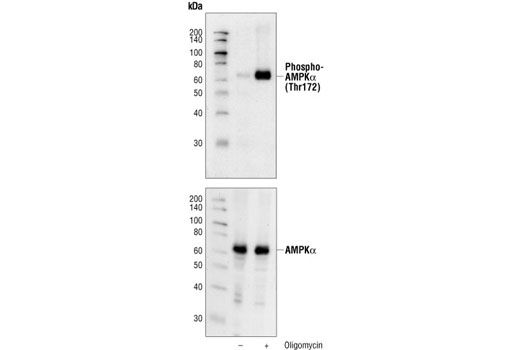
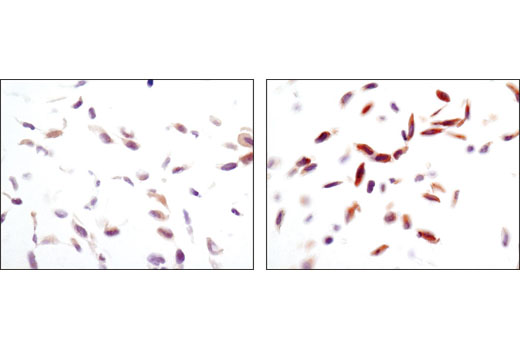
Monoclonal antibody is produced by immunizing animals with a synthetic peptide corresponding to residues surrounding Thr172 of human AMPKα protein.
AMP-activated protein kinase (AMPK) is highly conserved from yeast to plants and animals and plays a key role in the regulation of energy homeostasis (1). AMPK is a heterotrimeric complex composed of a catalytic α subunit and regulatory β and γ subunits, each of which is encoded by two or three distinct genes (α1, 2; β1, 2; γ1, 2, 3) (2). The kinase is activated by an elevated AMP/ATP ratio due to cellular and environmental stress, such as heat shock, hypoxia, and ischemia (1). The tumor suppressor LKB1, in association with accessory proteins STRAD and MO25, phosphorylates AMPKα at Thr172 in the activation loop, and this phosphorylation is required for AMPK activation (3-5). AMPKα is also phosphorylated at Thr258 and Ser485 (for α1; Ser491 for α2). The upstream kinase and the biological significance of these phosphorylation events have yet to be elucidated (6). The β1 subunit is post-translationally modified by myristoylation and multi-site phosphorylation including Ser24/25, Ser96, Ser101, Ser108, and Ser182 (6,7). Phosphorylation at Ser108 of the β1 subunit seems to be required for AMPK activation, while phosphorylation at Ser24/25 and Ser182 affects AMPK localization (7). Several mutations in AMPKγ subunits have been identified, most of which are located in the putative AMP/ATP binding sites (CBS or Bateman domains). Mutations at these sites lead to reduction of AMPK activity and cause glycogen accumulation in heart or skeletal muscle (1,2). Accumulating evidence indicates that AMPK not only regulates the metabolism of fatty acids and glycogen, but also modulates protein synthesis and cell growth through EF2 and TSC2/mTOR pathways, as well as blood flow via eNOS/nNOS (1).
Explore pathways related to this product.
STRING - Known and Predicted Protein-Protein Interactions.
Except as otherwise expressly agreed in a writing signed by a legally authorized representative of CST, the following terms apply to Products provided by CST, its affiliates or its distributors. Any Customer's terms and conditions that are in addition to, or different from, those contained herein, unless separately accepted in writing by a legally authorized representative of CST, are rejected and are of no force or effect.
Products are labeled with For Research Use Only or a similar labeling statement and have not been approved, cleared, or licensed by the FDA or other regulatory foreign or domestic entity, for any purpose. Customer shall not use any Product for any diagnostic or therapeutic purpose, or otherwise in any manner that conflicts with its labeling statement. Products sold or licensed by CST are provided for Customer as the end-user and solely for research and development uses. Any use of Product for diagnostic, prophylactic or therapeutic purposes, or any purchase of Product for resale (alone or as a component) or other commercial purpose, requires a separate license from CST. Customer shall (a) not sell, license, loan, donate or otherwise transfer or make available any Product to any third party, whether alone or in combination with other materials, or use the Products to manufacture any commercial products, (b) not copy, modify, reverse engineer, decompile, disassemble or otherwise attempt to discover the underlying structure or technology of the Products, or use the Products for the purpose of developing any products or services that would compete with CST products or services, (c) not alter or remove from the Products any trademarks, trade names, logos, patent or copyright notices or markings, (d) use the Products solely in accordance with CST Product Terms of Sale and any applicable documentation, and (e) comply with any license, terms of service or similar agreement with respect to any third party products or services used by Customer in connection with the Products.